Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.2.3.1.2 Specific (Acquired) Immune Response
The specific-response branch of the immune system is “adaptive” in the sense that the human body has the ability to learn to recognize novel foreign materials that it has never encountered before. The adaptive response increases in speed, in magnitude, and in defensive capabilities, after repeated exposure to a particular macromolecule. The specific-response branch of the immune system has two major components – the “humoral” and the “cell-mediated” responses.
The first component of the specific-response branch, also known as the humoral response, is the specialized white blood cells called bone marrow-derived cells, B lymphocytes, or B cells. Each of the ~0.1-1 trillion B cells usually present in the human body [352, 1756] displays on its surface just one type of the ~108-109 possible antigen receptor types [353, 354, 1764, 1765]. (~107 different specificities are thought to exist in a single individual [1756], but combinatorial human antibody libraries approaching ~1011 in diversity have been created [1766].) Each B cell makes a different receptor protein, so each B cell recognizes a different foreign molecule. Each B cell has ~105 membrane-bound antibody molecules which correspond to the one specific antibody that this cell is programmed to produce [1436]. If a B cell detects an intruder via a binding event to its particular receptor type, then that B cell divides rapidly, making a large number of clone cells bearing the same one unique receptor type. These cloned B cells then differentiate into plasma cells (cell lifetime ~1 week [1436]) that secrete free versions of the detected receptor type, also known as immunoglobulins (denoted “Ig”; Section 15.2.3.3) or, more commonly, antibodies,* into the bloodstream. These free antibodies bind to the surface of intruder cells. This makes the intruders visible to the innate system and allows them to be destroyed by the nonspecific complement/phagocyte system. There are five distinct classes of immunoglobulins – IgG, IgA, IgM, IgD, and IgE – representing ~25% of all human plasma proteins (Appendix B). Clones of the activated B cell line differentiate into memory B cells [356-359], giving rise to germinal centers (specialized structures within lymphoid organs) which allow a more rapid response if the same antigen is ever encountered again.
* “Natural” antibodies, produced spontaneously even in organisms raised in sterile environments, are often considered as part of the innate immune system [785, 786] and may serve a pathogen-herding function [787]. They usually have lower affinity and functional activity than antibodies developed as part of a specific immune response.
The second component of the specific-response branch, also known as the cell-mediated response, is the specialized white blood cells called thymus-derived cells, T lymphocytes, or T cells. T cells serve a variety of purposes, such as recognizing and killing cells bearing nonself molecules on their surface, or helping B cells produce antibodies. Both B and T lymphocytes range from 6-12 microns in diameter [5334-5339]. There can be as many as ~1 trillion T cells in the body [353, 360]. To maximize the chances of encountering antigen, lymphocytes recirculate from blood to lymphoid tissues (Section 8.2.1.3) and back dozens of times per day – the ~1010 lymphocytes in human blood have a mean residence time of ~30 minutes in the blood [1763]. Each T cell has ~105 molecules of a specific antigen-binding T cell receptor exposed on its surface [1436]. The term CD or “cluster of differentiation” is used to designate surface marker proteins that distinguish various T cell subpopulations – including most importantly the CD4 lymphocytes (mature helper-inducer T cells) and the CD8 lymphocytes (cytotoxic/killer and suppressor T cells). The CD3 polypeptide complex is expressed on the surface of T cells and thymocytes. About 75% of all peripheral blood lymphocytes of normal individuals are T cells (i.e., ~25% are B cells), of which ~50% are helper-inducer CD4 T cells and ~25% are suppressor or cytotoxic CD8 T cells [1756]. Many T cell functions are mediated by cytokines (Table 7.2), most notably the interleukins (IL), interferons (IFN), tumor necrosis factor (TNF), and granulocyte-macrophage colony stimulating factor (GM-CSF). Surface markers and life span of a population of human T cells equivalent to murine memory T cells have been demonstrated [362, 363], thus the existence of memory T cells in humans, though once controversial [364], is now well established [365-370].
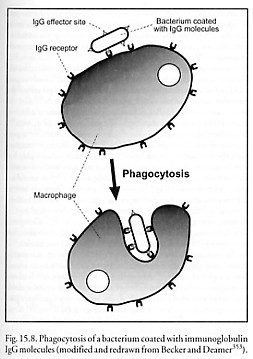
The immune system employs two systems of recognition elements – soluble antibodies and cell-attached T-cell receptors. Both are produced by similar types of genetic elements (the immunoglobulin gene superfamily). Opsonization is the process by which soluble antibodies bind to and coat antigenic particles to mediate destruction by phagocytosis (Figure 15.8) or complement activation [5491], and can also block the action of proteins required for pathogen invasiveness. Cell-bound T-cell receptors (having binding sites biochemically similar to antibodies) provide protection against intracellular microbes such as viruses and mycobacteria. Such microbes are shielded from antibodies by the host cell membrane. But nearly all vertebrate cell types exhibit a sample of peptides on their surface, derived from the digestion of proteins in their cytosol, which includes fragments of microbe or virus proteins, if any are present. T cells use their T-cell receptors to continually scan the surfaces of cells in order to detect, and then kill, any cells displaying recognized foreign markings.
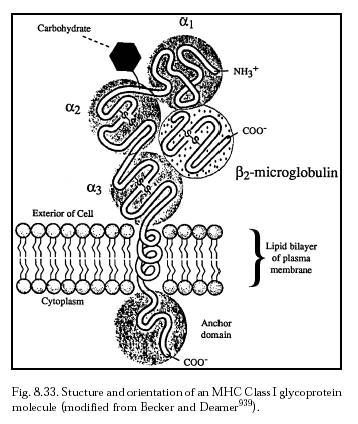
Both humoral and cell-mediated immune responses require a signaling step that announces the presence of non-self protein. In most cases this signaling step is the end result of a sequence of intracellular processes during which a foreign protein is broken down and processed into short oligopeptides, typically 8-10 amino acids long. Inside the cell, these antigenic fragments then become associated with a set of glycoproteins called the major histocompatibility complex (MHC) molecules (Section 8.5.2.1). The function of the MHC molecules is to pick up the peptide fragments and convey them to the cell membrane for the purpose of T-cell recognition and proliferation, a process called antigen presentation. (Autoimmune disease results when self-proteins presented on the MHC are misrecognized as non-self.)
Peptide fragments derived from endogenous intracellular microbes are transported to the outside of the cell complexed with MHC class I molecules (Figure 8.33) where they can then present antigen to CD8 lymphocytes. Most cells have MHC-I molecules and the constant sampling of the cytosolic protein population provides a powerful surveillance mechanism geared to detecting viral infection or intracellular bacteria. Protection against intracellular microbes is largely the domain of cell-mediated immunity, although for several pathogens antibody responses also contribute to host defense [1760]. It would seem that, from the point of view of avoiding immune attacks on cells in which intracellular cell repair machines are present, it will generally be important to avoid allowing these machines to be swept along on the MHC antigen presentation machinery and presented on the surface of cells.
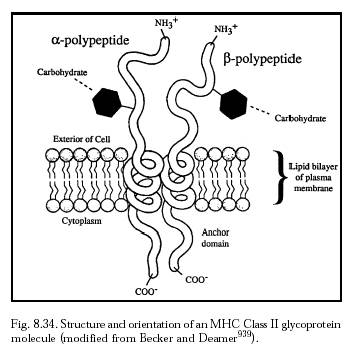
Peptide fragments derived from exogenous antigens (either free or complexed with antibody) that have been endocytosed into the cell are transported to the cell surface and presented to CD4 cells by MHC class II molecules (Figure 8.34). Class II molecules occur only in specialized cell types called antigen presenting cells (APCs) such as B cells, macrophages, and dendritic cells. Peptides presented by MHC II molecules can activate the appropriate cascade of chemical processes that lead to antibody formation.* Of course, intracellular nanorobots could load the local MHC population with artifactual or manufactured antigens to manipulate the immune system response.
* Superantigens are polypeptide toxins secreted by bacteria [2243] or produced by viruses [2244] that interfere with normal immune function by directly activating T cells in a nonspecific manner. Superantigens are thought to induce aberrant cell-cell interactions by binding simultaneously to T cell antigen receptors and to MHC class II molecules containing certain relatively rare peptide fragments [2238-2241]. They are believed to be associated with toxic shock [2241, 2242], food poisoning [2245], and various autoimmune disorders [2246].
Dendritic cells (DCs) are antigen presenting cells scattered throughout the body, including the various portals of microbial entry, where they reside in immature form as immunological sensors [2555]. They are leukocytes of bone marrow origin [5342]. Like lymphocytes, DCs form subsets differing in phenotype, function, and locale. In human skin, DCs called Langerhans cells [5340-5342] are found in the epidermis. Interstitial DCs are found in the dermis. In human blood, one type of immature DC differentiates into mature DCs in response to inflammatory stimuli, and another precursor DC responds to viruses, tumors, and other stimuli by differentiating into mature plasmacytoid DCs [2556-2559]. In mice, other DCs have also been found in the thymic cortex and T cell areas of secondary lymphoid organs, in the marginal zones of the spleen, in the subcapsular sinuses of the lymph nodes, and in the subepithelial dome of Peyer’s patches (oval masses of lymphoid tissue on the mucous membrane lining the small intestine) [2557-2559].
When a microbe invades a tissue, resident immature DCs employ pattern recognition receptors [2560-2565] embedded in their plasma membrane surface to sense the microbe by recognizing evolutionarily conserved molecular patterns that are integral to microbial carbohydrates, lipids, and nucleic acids [2555]. For example, lipopolysaccharide (LPS) from E. coli is recognized by Toll-like receptor TLR4; peptidoglycans from Staphylococcus aureus and LPS from Porphyromonas gingivalis are recognized by TLR2; bacterial flagellin is recognized by TLR5; and so forth [2561-2564]. Once a specific microbe is detected, physical information about the pathogen is captured via phagocytosis, endocytosis, or micropinocytosis for soluble antigens [2557] and is relayed to naive T lymphocytes in the draining lymph nodes. Immature DCs exit the site of infection and migrate toward the T cell areas of the proximal lymph nodes via the afferent lymphatics, following rising gradients of 6Ckine and MIP-3beta cytokines expressed in lymphoid organs [2557]. During this journey the DCs mature, losing their antigen-capturing capacities but acquiring the ability to process and display peptide antigens on their surface via MHC class II molecules for antigen presentation to T-lymphocytes, as previously described.
Last updated on 30 April 2004
{kind=link}
{kind=link}
{kind=link}
{kind=link}