Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.5.7.3.1 Mechanical Cytoskeleton Disorganization
Generalized disruption of the cytoskeleton can be very harmful to living cells. Disorganization of the cytoskeletal architecture has been associated with diseases as diverse as heart failure [4310, 4311], rotavirus infection [4312], sickle cell anemia [5676-5678], lissencephaly [5673], and Alzheimer’s disease [4313-4315, 5682]. A “collapse transition” of neurofilament sidearm domains may contribute to amyotrophic lateral sclerosis (ALS) and Parkinson’s disease [5679-5681]. Stress-related cytoskeletal fracture can be caused by 1-Hz stresses imposed by a mechanical probe on isolated rat ventricular myocytes [3858]. Cancer cells forced through 5- to 12-micron pores in polycarbonate membrane suffer traumatic spatial dissociation between components of the cell periphery, the cytoskeleton, and nucleus, inducing a ~1-week dormant state in the cells due to the mechanical trauma [4316].
Nanorobots could induce various cell pathologies by mechanically disrupting specialized cytoskeletons consisting of cytoplasmic networks of ~6-nm diameter actin microfilaments, ~10-nm intermediate filaments, ~25-nm microtubules, or their many associated proteins [4317, 4334] (Section 8.5.3.11), with effects similar to those of chemical disruption [4318]. Functions of these specialized cytoskeletons that could be disturbed include mechanical integrity and wound-healing in epidermal cells, cell polarity in simple epithelia, contraction in muscle cells, hearing and balance in the inner ear cells, axonal transport in neurons, and neuromuscular junction formation between muscle cells and motor neurons [4334].
As a nanorobot enters the cell, the first risk is transmembrane linkage disruptions. Muscular dystrophy may be caused by disorganization of links between the intracellular cytoskeleton and the ECM [4003], and the disruption of proper adhesive interactions with neighboring cells can lead to fatal defects in extracellular tissue architecture [4319]. Epithelial cells subjected to mechanical strain may release in vivo proteases to cut intercellular adhesions [4281]. Looking inward, cellular mechanoprotective adaptations involve a coordinated remodeling of the cell membrane and the associated cytoskeleton [4320]. For example, the breakage of major cytoskeletal attachments between the plasma membrane and peripheral myofibers in cardiac myocytes predisposes the cell to further mechanical damage from cell swelling or from ischemic contracture [4321]. As another example, when infecting parsley cells, attacking fungus cells extend a penetrating hypha through the cell membrane, eliciting a defensive cytoskeletal reorganization [4322]. A local mechanical stimulus produced by a needle of the same diameter as the fungal hypha inserted through the host cell wall similarly induces the translocation of cytoplasm and nucleus to the site of stimulation, the generation of intracellular reactive oxygen intermediaries, and the expression of some elicitor-responsive genes. Without the mechanical stimulation, the morphological changes are not detected [4322].
Mechanical disruption of cytoskeleton associated proteins by passing nanorobots could produce various cytopathologies. For instance, plectin is a 580 kD intracellular protein that links intermediate filaments with actin microfilaments, microtubules, and plasma membrane. Disruption of plectin function results in severe skin blistering and muscular degeneration, consistent with plectin’s role in stabilizing cells against external mechanical forces [4323] and as a regulator of intracellular actin dynamics [4324]. Disturbance of centrosomes [4325, 4326] or other in cyto fixed polarity markers [4327] could result in developmental or morphogenetic defects during subsequent cell division. Mechanical disturbance of cytoskeleton associated proteins could also alter the mechanical properties of cells, such as the ability of the cytoskeleton to deform and flow. One research group [4649] believes cells exist close to a glass transition state, a state regulated by cytoskeletal proteins which modulate the effective noise temperature of the matrix; this state thus could also, in principle, by manipulated by nanorobots.
Actin microfilaments might be disrupted by the mechanical activities of medical nanorobots. In the simplest case, endothelium exposed to shear stress undergoes cell shape change, alignment, and microfilament network remodeling in the direction of flow, though these changes can be blocked with nocodazole [3803]. Glomerular distention is also associated with cellular mechanical strain. A contractile cytoskeleton in mesangial cells, formed by F-actin-containing stress fibers, maintains structural integrity and modulates glomerular expansion. Mesangial cells have a cytoskeleton capable of contraction that is disorganized in long-term diabetes. Disorganization of stress fibers may be a cause of hyperfiltration in diabetes [4328]. Cutting the actin lattice may diminish both cell contractility [4329] and mechanical signal transduction into the nucleus [4330]. Care must also be taken to ensure that the surfaces or activities of intracellular nanorobots do not provide unplanned foci for actin polymerization, given that the kinetics of actin polymerization is autocatalytic and that the actin-based motility of functionalized microspheres can be reconstituted in vitro from only five pure proteins [4331]. Widespread actin disruption might produce symptoms analogous to elliptocytosis [5398-5403] and other inherited hemolytic disorders [5405-5408] that are caused by disorganization of the subsurface spectrin-actin cell cortex in the erythrocyte [5404].
Intermediate filaments might also be disrupted mechanically. Perturbations in the architecture of the intermediate filament cytoskeleton in keratinocytes and in neurons can lead to degenerative diseases of the skin, muscle cells, and nervous system [4332-4336]. Knockout of the extensive keratin filament network jeopardizes the mechanical integrity of the epidermal cell, producing cell fragility and cytolysis manifesting as blistering skin disorders [4332]. Tissues lacking intermediate filaments fall apart, are mechanically unstable, and cannot resist physical stress, which leads to cell degeneration [4337]. Perinuclear clumping of fragmented keratin intermediate filaments accompanies many keratin disorders of skin, hair, and nails [4338]. In active muscle, intermediate filaments play an important role in the organization and stabilization of myofibril-membrane attachment sites. Their disruption can eliminate the deep membrane invaginations that are normally present in the healthy sarcolemma [4333]. Neuronal intermediate filaments are normally anchored to actin cytoskeleton. If this anchoring fails, the cell displays short, disorganized and unstable microtubules that are defective in axonal transport. Neuronal survival requires viable interconnects among all three cytoskeletal networks [4336]. Impairment of normal axonal cytoskeletal organization in Charcot-Marie-Tooth disease results in distal axonal degeneration and fiber loss [4339].
The microtubule cytoskeleton could become disorganized due to careless or intentional intracellular operations by nanorobots, possibly: (1) simulating congenital brain malformation [4340]; (2) giving results similar to treatment with vincristine, a microtubule depolymerizing drug that produces peripheral neuropathy in humans accompanied by painful paresthesias and dysesthesias [4341]; or (3) giving results similar to treatment with ethanol, leading to oxidative injury producing a loss of gastrointestinal barrier integrity [4342]. Mechanical disturbance of the microtubule cytoskeleton induces electrophysiological modification of cell-cycle-dependent EAG potassium channels in mammalian tissue cells [4343], and mechanical strain can induce a major decline in tubulin production in osteoblasts [4344]. Nanorobot mechanical operations could also induce buckling and loop formation of tubulin fibers, as has been observed [5674] inside shrinking vesicles when the surface tension of the shrinking bubble overcomes the Euler buckling strength of the fibers; intracellular tubulin twisted into 5-micron tennis-racquet shapes has also been observed [5675].
Microtubules allowed to form under microgravity conditions show almost no self-organization and are locally disordered, unlike microtubules formed in 1-g conditions [4345]. Nanorobotic manipulations of cytoskeletal elements that offset, reduce, or cancel the stimulative effects of normal gravity could produce the same sort of cellular architectural disorganization as observed under microgravity conditions (Section 4.4.2 and Chapter 28) that alters the pattern of microtubular orientation [4346].
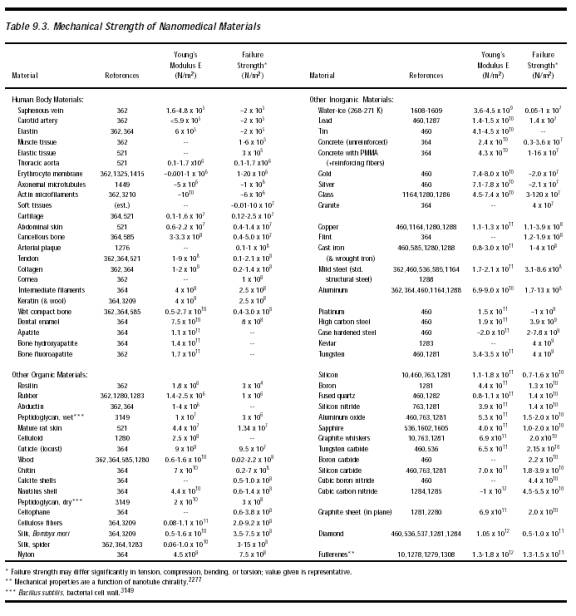
A nanorobot with sharp edges that cuts a microtubule probably cleaves the hydrogen bonds between the alpha and beta monomers, rather than the covalent bonds within the monomers. This creates a new “plus” and “minus” end for the microtubule. In most cases this would not be fatal for the cell and in fact normally would have little impact because large-scale microtubule network patterns (e.g., asters, whorls, and interconnected pole networks) are self-assembling and are motor-molecule concentration-dependent [4347]. Nevertheless, in cyto nanorobots should avoid physically severing cytoskeletal elements whenever possible. Simple estimates of mechanical strength (Table 9.3) applied to typical fiber diameters suggest that the tensile failure strengths are ~170 pN for actin microfilaments, ~300-500 pN for microtubules and ~20,000 pN for intermediate filaments. Nanorobots should avoid applying local forces of these magnitudes or larger in the vicinity of such fibers.
Force thresholds for cellular activation (Section 15.5.4.1) may be considerably less than the indicated tensile failure strengths. In 2002, the absolute force thresholds for failure, the range of mechanical frequency responses, and the threshold fraction of disturbed cytoskeleton required to elicit cellular response all had yet to be precisely determined. For example, during mitosis a force of 15-20 pN is required to detach microtubule-bound chromosomes [4350] but a tensile force of up to 210 pN is required to detach a microtubule from a kinetochore [4348]. Moreover, a nanorobot presenting a 1-micron2 forward surface during intracellular locomotion through a (20 micron)3 tissue cell intercepts only ~0.25% of the entire cytoskeleton during each 20-micron of transcellular travel. In cyto medical nanorobots may be restricted to speeds of ~10 microns/sec while traversing intracellular clear paths (Section 8.5.3.12) and ~1 micron/sec during transfilamentary intracellular locomotion, with progressive resealing of cytoskeletal elements that must be temporarily severed to allow the nanorobot to pass (Section 9.4.6). Intranuclear locomotion conservatively should progress no faster than natural chromosomal dragging rates during mitosis [4349, 4350], or ~0.1 micron/sec, applying forces of at most ~50 pN (Section 9.4.6).
Last updated on 30 April 2004
{kind=link}